|
VHJ van der Velden, PJM Leenen & HA Drexhage
Department of Immunology, Erasmus University, Rotterdam, The Netherlands
Correspondence to: VHJ van der Velden, Department of Immunology, Erasmus University Rotterdam, PO Box 1738, 3000 DR Rotterdam, The Netherlands; Fax: +31 10 408 9456
|
TO THE EDITOR Recently it has been shown that CD13/aminopeptidase N (APN) is involved in the development of dendritic cells (DC) and macrophages from cord blood CD34+ cells.1 More specifically, CD13 expression was positively correlated with cell proliferation, and inhibition of CD13/APN resulted in apoptosis. Previously, we have reported expression of CD13/APN by DC in the human lung,2 as well as by various populations of mouse DC.3,4 Given the implication of CD13/APN activity in a variety of cellular functions and the prominent expression by DC, we approached the question of whether CD13/APN is functionally involved with DC maturation. To test this, we cultured purified human monocytes in culture fluid with granulocyte–monocyte colony-stimulating factor (GM-CSF) and interleukin (IL)-4 for 5 days, resulting in the development of CD1a-positive immature DC.5 These cultures were performed in the absence or presence of CD13/APN inhibitors: the potent and CD13/APN-specific inhibitor leuhistin and the less potent inhibitor bestatin (both 10-3 M). As an additional control, monocytes were cultured for 5 days in culture fluid without GM-CSF and IL-4. Subsequently, the cells were immunophenotyped by flow cytometry and viability of the cells was determined by propidium iodide exclusion. After 5 days of culture with GM-CSF and IL-4, in either in or not in the presence of CD13/APN inhibitors, two main cell populations could be distinguished on scatter characteristics (in addition to contaminating lymphocytes): one population with high forward and side scatter and one population with low forward scatter and low/intermediate side scatter (Figure 1a). Using propidium iodide it appeared that the population with high scatter characteristics comprised mainly viable cells (>80%), whereas the population with low forward scatter had a low viability (<30%). In the absence of CD13/APN inhibitors, the high scatter population comprised approximately 50% of the cells (Figure 1b). Immunophenotyping of this population showed that these cells expressed CD1a, HLA-DR, CD13, and CD80, but almost lacked CD14 expression (Figure 2a), which is consistent with the development of immature DC. Interestingly, when leuhistin was added to the culture, the development of immature DC was significantly inhibited and this was accompanied by an increase in the low scatter population, ie dead cells (Figure 1b). It is unlikely that this is due to nonspecific cytotoxicity of leuhistin, as contaminating lymphocytes were not affected. Addition of bestatin during the DC culture did not affect the relative composition of the two main cell populations (Figure 1b). As such, this finding of increased cellular death induced by blockade of CD13/APN by potent inhibitors is in accordance with the observation that inhibition of CD13/APN resulted in apoptosis.1 Addition of leuhistin to the DC culture, however, not only affected the development of the high scatter population, but also affected the immunophenotype of these cells as compared to cells cultured without CD13/APN inhibitors. Addition of leuhistin strongly reduced the percentage CD1a and CD80 expressing cells, which was accompanied by a strong reduction in fluorescence intensity of the positive cells. In contrast, the percentage of CD14 expressing cells was significantly increased (Figure 2). The immunophenotype of these cells resembled the immunophenotype of monocytes cultured in culture fluid without the maturation-inducing factors GM-CSF and IL-4. These cells expressed CD14, HLA-DR, CD13, and low levels of CD80, but did not express CD1a (Figure 2). Bestatin had similar effects as leuhistin (Figure 2), although these were less pronounced, possible due to its lower potency. Under all culture conditions, CD13/APN was present on 100% of the large scatter population, but the level of expression was strongly increased when APN inhibitors were added to the culture (Figure 2). This enhanced expression may reflect a cellular mechanism to circumvent the inhibition of CD13. Overall, the above data indicate that inhibition of CD13/APN prevented the development of immature DC and maintained the cells in a monocyte-like stage. This is further supported by our preliminary finding that addition of CD13/APN inhibitors during the culture of DC from monocytes also significantly reduces the capacity of the resulting cells to stimulate mixed leukocyte reactions. In conclusion, our results show that CD13/APN plays a role in the development of (immature) DC from monocytes. In addition to its reported effect on cell viability by preventing increased apoptosis induction,1 we now demonstrate that CD13 also affects the maturation of the DC. The molecular basis of this effect is unclear. Hypothetically, CD13/APN may be involved with the utilization of GM-CSF by the cells, as suggested by Rosenzwajg et al.1 An alternative, but similarly hypothetical explanation is that CD13/APN activates autocrine or paracrine maturation-promoting peptides. In our view, these are interesting options to be tested given the potent effects of CD13/APN inhibitors on myeloid cell growth and DC maturation.
|
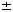 s.e.m.; n = 3). Total absolute cell numbers did not differ significantly among the different culture conditions.
s.e.m.; n = 3). Total absolute cell numbers did not differ significantly among the different culture conditions.